Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review

Abstract
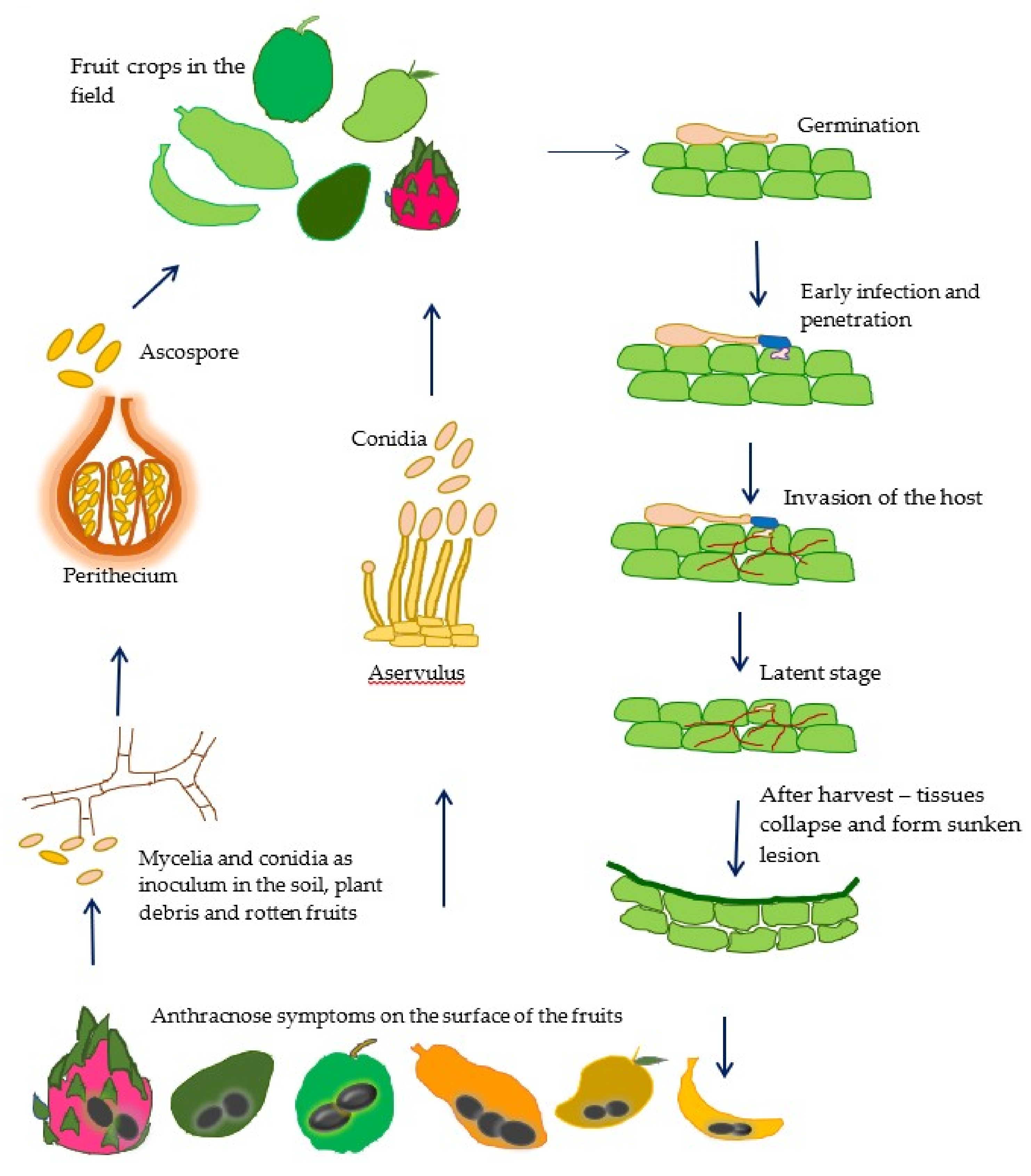
1. Introduction
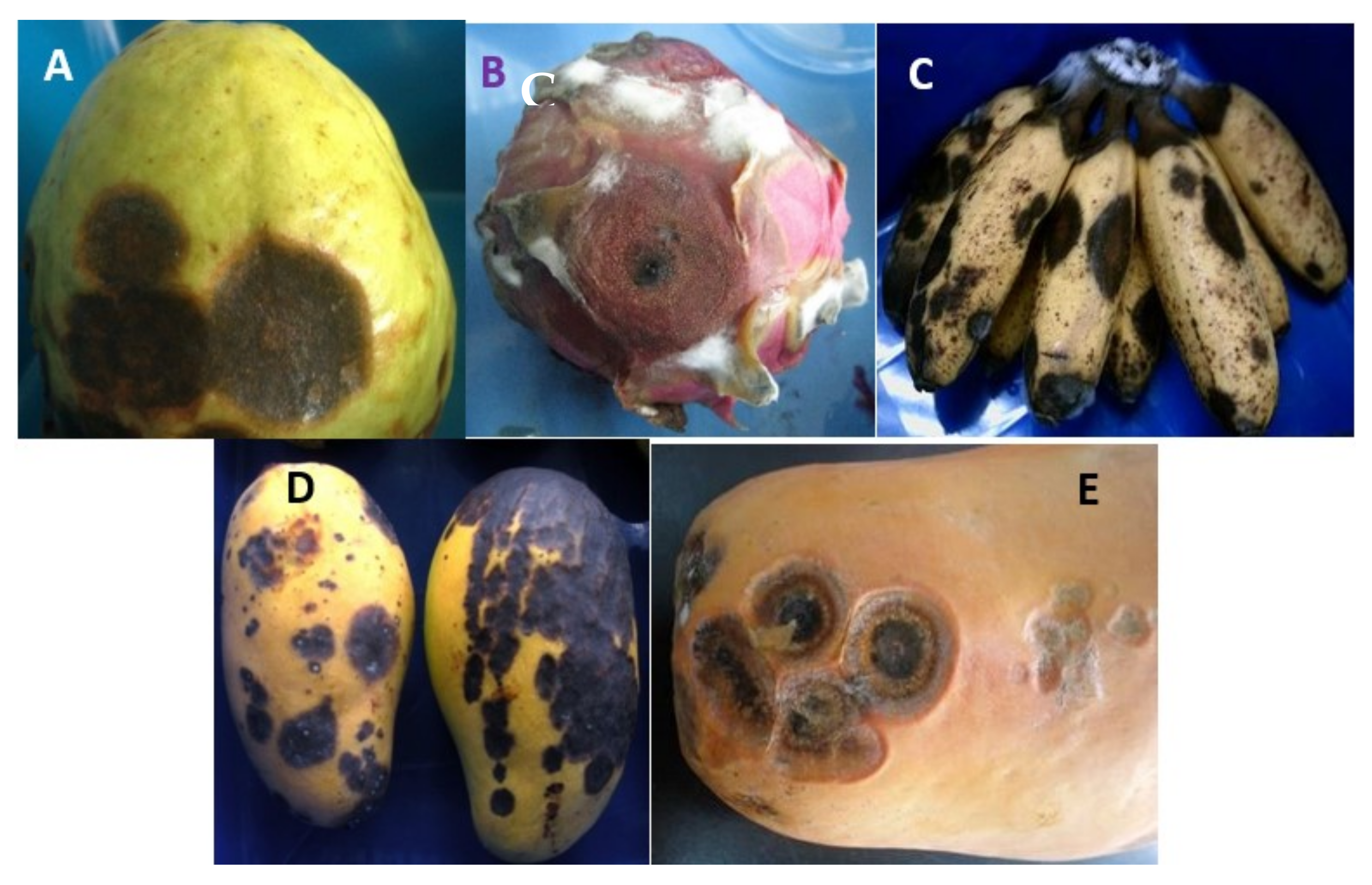
2. Banana Anthracnose
3. Mango Anthracnose
4. Papaya Anthracnose
5. Dragon Fruit Anthracnose
6. Guava Anthracnose
7. Avocado Anthracnose
8. Present and Future Management of Anthracnose
9. Conclusions and Future Directions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| ACT | actin |
| ApMat | mating type (Mat1-2) |
| CHS1 | chitin synthase |
| CAL | calmodulin |
| CFU | colony forming unit |
| GAPDH | glyceraldehyde-3-phosphatedehydrogenase |
| GRAS | generally recognized as safe |
| GS | glutamine synthetase |
| HIS3 | histone |
| ITS | internal transcribed spacer |
References
- OECD; FAO. OECD–FAO Agricultural Outlook 2019–2028Part. II—OverviewSpecial Feature and Commodity Chapters; OECD Publishing: ParisFrance2019. [Google Scholar]
- AltendorfS. Minor. Tropical Fruits: Mainstreaming a Niche Market; Food and Agriculture Organization of the United Nations: RomeItaly2018; pp. 69–76. [Google Scholar]
- OECD; FAO. OECD–FAO Agricultural Outlook 2020–2029; FAO: RomeItaly; OECD Publishing: ParisFrance2020. [Google Scholar]
- AltendorfS. Major Tropical Fruits Statistical Compendium 2018; FAO: RomeItaly2019. [Google Scholar]
- FreemanS.; KatanT.; ShabiE. Characterization of Colletotrichum species responsible for anthracnose diseases of various fruits. Plant Dis. 199882596–605. [Google Scholar] [CrossRef]
- PruskyD. Pathogen quiescence in postharvest diseases. Annu. Rev. Phytopathol. 199634413–434. [Google Scholar] [CrossRef] [PubMed]
- JeffriesP.; DoddJ.C.; JegerM.J.; PlumbleyR.A. The biology and control of Colletotrichum species on tropical fruit crops. Plant Pathol. 199039343–366. [Google Scholar] [CrossRef]
- WhartonP.S.; Dieguez-UribeondoJ. The biology of Colletotrichum acutatum. An. Jardín Botánico Madr. 2004613–22. [Google Scholar]
- TalhinhasP.; Mota-CapitaoC.; MartinsS.; RamosA.P.; Neves-MartinsJ.; Guerra-GuimaraesL.; VarzeaV.; SilvaM.C.; SreenivasaprasadS.; OliveiraH. Epidemiologyhistopathology and aetiology of olive anthracnose caused by Colletotrichum acutatum and C. gloeosporioides in Portugal. Plant Pathol. 201160483–495. [Google Scholar] [CrossRef]
- ShawM.W.; EmmanuelC.J.; EmildaD.; TerhemR.B.; ShafiaA.; TsamaidiD.; EmblowM.; VanK.; JanA.L. Analysis of cryptic systemic Botrytis infections in symptomless hosts. Front. Plant Sci. 201671–14. [Google Scholar] [CrossRef]
- FitzellR.D.; PeakC.M. The epidemiology of anthracnose disease of mango: Inoculum sourcesspore production and dispersal. Ann. Appl. Biol. 198410453–59. [Google Scholar] [CrossRef]
- DammU.; CannonP.F.; WoudenbergJ.H.C.; CrousP.W. The Colletotrichum acutatum species complex. Stud. Mycol. 20127337–113. [Google Scholar] [CrossRef]
- WeirB.; JohnstonP.R.; DammU. The Colletotrichum gloeosporioides species complex. Stud. Mycol. 201273115–180. [Google Scholar] [CrossRef]
- CannonP.F.; DammU.; JohnstonP.R.; WeirB.S. Colletotrichum—Current status and future directions. Stud. Mycol. 201273181–213. [Google Scholar] [CrossRef]
- De SilvaD.D.; CrousP.W.; AdesP.K.; HydeK.D.; PaulP.W.J. Lifes of Colletotrichum species and implications for plant biosecurity. Fungal Biol. Rev. 201731155–168. [Google Scholar] [CrossRef]
- PhoulivongS.; McKenzieE.; HydeK. Cross infection of Colletotrichum species; a case study with tropical fruits. Curr. Res. Environ. Appl. Mycol. 2012299–111. [Google Scholar] [CrossRef]
- PengL.J.; SunT.; YangY.L.; CaiL.; HydeK.D.; BahkaliA.H.; LiuZ.Y. Colletotrichum species on grape in Guizhou and Yunnan provincesChina. Mycoscience 20135429–41. [Google Scholar] [CrossRef]
- FreemanS.; HorowitzS.; SharonA. Pathogenic and nonpathogenic lifes in Colletotrichum acutatum from strawberry and other plants. Phytopathology 200191986–992. [Google Scholar] [CrossRef]
- SandersG.M.; KorstenL. A comparative morphological study of South African avocado and mango isolates of Colletotrichum gloeosporioides. Can. J. Bot. 200381877–885. [Google Scholar] [CrossRef]
- Intan SakinahM.A.; SuziantiI.V.; LatiffahZ. First report of Colletotrichum gloeosporioides causing anthracnose of banana (Musa spp.) in Malaysia. Plant Dis. 201397991. [Google Scholar] [CrossRef] [PubMed]
- ZhouY.; HuangJ.S.; YangL.Y.; WangG.F.; LiJ.Q. First report of banana anthracnose caused by Colletotrichum scovillei in China. Plant Dis. 2016101381. [Google Scholar] [CrossRef]
- KumarV.S.; NairB.A.; NairP.V.R.; AnnamalaiA.; JaishankerR.; UmamaheswaranK.; SoorajN.P.; PeethambaranC.K. First report of Colletotrichum siamense causing anthracnose of cliff Banana in India. Plant Dis. 2017101390. [Google Scholar] [CrossRef]
- VieiraW.A.S.; LimaW.G.; NascimentoE.S.; MichereffS.J.; CâmaraM.P.S.; DoyleV.P. The impact of phenotypic and molecular data on the inference of Colletotrichum diversity associated with Musa. Mycologia 2017109912–934. [Google Scholar] [CrossRef] [PubMed]
- RieraN.; Ramirez-VillacisD.; Barriga-MedinaN.; Alvarez-SantanaJ.; HerreraK.; RualesC.; Leon-ReyesA. First report of banana anthracnose caused by Colletotrichum gloeosporioides in Ecuador. Plant Dis. 2019103763. [Google Scholar] [CrossRef]
- UysalA.; KurtS. First report of fruit and leaf anthracnose caused by Colletotrichum karstii on avocado in Turkey. Crop. Prot. 2020133105145. [Google Scholar] [CrossRef]
- Fuentes-AragónD.; Rebollar-AlviterA.; Osnaya-GonzálezM.; Enciso-MaldonadoG.A.; González-ReyesH.; Silva-RojasH.V. Multilocus phylogenetic analyses suggest the presence of Colletotrichum chrysophilum causing banana anthracnose in Mexico. J. Plant Dis. Prot. 20201–7. [Google Scholar] [CrossRef]
- RojasE.I.; RehnerS.A.; SamuelsG.J.; Van BaelS.A.; HerreE.A.; CannonA.; ChenR.; PangJ.; WangR.; ZhangY.; et al. Colletotrichum gloeosporioides s.l. associated with Theobroma cacao and other plants in Panama: Multilocus phylogenies distinguish host-associated pathogens from asymptomatic endophytes. Mycologia 20101021318–1338. [Google Scholar] [CrossRef] [PubMed]
- LimaN.B.; BatistaM.V.A.; Morais JuniorM.A.; BarbosaM.A.G.; MichereffS.J.; HydeK.D.; CâmaraM.P.S. Five Colletotrichum species are responsible for mango anthracnose in northeastern Brazil. Fungal Divers. 20136175–88. [Google Scholar] [CrossRef]
- SharmaG.; KumarN.; WeirB.S.; HydeK.D.; ShenoyB.D. Apmat gene marker can resolve Colletotrichum species: A case study with Mangifera indica. Fungal Divers. 201361117–138. [Google Scholar] [CrossRef]
- KrishnapillaiN.; WijeratnamR.S.W. First report of Colletotrichum asianum causing anthracnose on Willard mangoes in Sri Lanka. New Dis. Rep. 201429. [Google Scholar] [CrossRef]
- LatiffahZ.; Nurul ZaadahJ.; SuziantiI.V.; Intan SakinahM.A. Molecular characterization of Colletotrichum isolates associated with anthracnose of mango fruit. Sains Malays. 201544651–656. [Google Scholar]
- Pardo-De la HozC.J.; CalderonC.; RinconA.M.; CardenasM.; DaniesG.; Lopez-Kleine RestrepoS.; JimenezP. Species from the Colletotrichum acutatumColletotrichum boninense and Colletotrichum gloeosporioides species complexes associated with tree tomato and mango crops in Colombia. Plant Pathol. 201665227–237. [Google Scholar] [CrossRef]
- QinL.P.; HuangS.L.; LinS.H. First report of anthracnose of Mangifera indica caused by Colletotrichum asianum in Sanya city in China. Plant Dis. 20171011038. [Google Scholar] [CrossRef]
- MoJ.; ZhaoG.; LiQ.; SolangiG.S.; TangL.; GuoT.; HuangS.; HsiangT. Identification and characterization of Colletotrichum species associated with mango anthracnose in GuangxiChina. Plant Dis. 20181021283–1289. [Google Scholar] [CrossRef]
- LiQ.; BuJ.; ShuJ.; YuZ.; TangL.; HuangS.; GuoT.; MoJ.; LuoS.; GhulamS.S.; et al. Colletotrichum species associated with mango in southern China. Sci. Rep. 2019918891. [Google Scholar] [CrossRef] [PubMed]
- WuC.; ChenH.; NiH. Identification and characterization of Colletotrichum species associated with mango anthracnose in Taiwan. Eur. J. Plant Pathol. 20201571–15. [Google Scholar] [CrossRef]
- Tovar-PedrazaJ.M.; Mora-AguileraJ.A.; Nava-DíazC.; LimaN.B.; MichereffS.J.; Sandoval-IslasJ.S.; CâmaraM.P.S.; Téliz-OrtizD.; Leyva-MirS.G. Distribution and pathogenicity of Colletotrichum species associated with mango anthracnose in Mexico. Plant Dis. 2020104137–146. [Google Scholar] [CrossRef]
- AlvarezL.V.; HattoriY.; DeocarisC.C.; MapanaoC.P.; BautistaA.B.; CanoM.J.B.; NaitoK.; KitabataS.; MotohashiK.; NakashimC. Colletotrichum asianum causes anthracnose in Philippine mango cv. CarabaoAustralas. Plant Dis. Notes 20201513. [Google Scholar] [CrossRef]
- AhmadT.; WangJ.; ZhengY.; MugiziA.E.; MoosaA.; ChengrongN.; LiuY. First record of Colletotrichum alienum causing postharvest anthracnose disease of mango fruit in China. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Tapia-TussellR.; Quijano-RamayoA.; Cortes-VelazquezA.; LappeP.; Larque-SaavedraA.; Perez-BritoD. PCR-based detection and characterization of the fungal pathogens Colletotrichum gloeosporioides and Colletotrichum capsici causing anthracnose in papaya (Carica papaya L.) in the Yucatan Peninsula. Mol. Biotech. 200840293–298. [Google Scholar] [CrossRef] [PubMed]
- RahmanM.A.; MahmudT.M.M.; KadirJ.; Abdul RahmanR.; BegumM.M. Major postharvest fungal diseases of papaya cv. ‘Sekaki’ in SelangorMalaysia. Pertanika J. Trop. Agric. Sci. 20083127–34. [Google Scholar]
- YaguchiY.; NakanishiY.; SaitoT.; NakamuraS. Anthracnose of Carica papaya L. caused by Colletotrichum capsici. Ann. Phytopath. Soc. Jpn. 199561222. [Google Scholar]
- TarnowskiT.B.L.; PloetzR.C. First report of Colletotrichum capsici causing postharvest anthracnose on papaya in South Florida. Plant Dis. 2010941065. [Google Scholar] [CrossRef]
- NascimentoR.J.; MizubutiE.S.G.; CâmaraM.P.S.; FerreiraM.F.; MaymonM.; FreemanS.; MichereffS.J. First report of papaya fruit rot caused by Colletotrichum magna in Brazil. Plant Dis. 2010941506. [Google Scholar] [CrossRef]
- Santamaría BasultoF.; Díaz PlazaR.; Gutiérrez AlonsoO.; Santamaría FernándezJ.; Larqué SaavedraA. Control of two species of Colletotrichum causing anthracnose in Maradol papaya fruits. Rev. Mex. Cienc. Agríc. 20172631–643. [Google Scholar]
- DammU.; CannonP.F.; WoudenbergJ.H.C.; JohnstonP.R.; WeirB.S.; TanY.P.; ShivasR.G.; CrousP.W. The Colletotrichum boninense species complex. Stud. Mycol. 2012731–36. [Google Scholar] [CrossRef]
- RampersadS.N. Molecular and phenotypic characterization of Colletotrichum species associated with anthracnose disease of papaya in Trinidad. Plant Dis. 2011951244–1254. [Google Scholar] [CrossRef]
- MaharajA.; RampersadS.N. Genetic Differentiation of Colletotrichum gloeosporioides and C. truncatum associated with anthracnose disease of papaya (Carica papaya L.) and bell pepper (Capsium annuum L.) based on ITS PCR-RFLP fingerprinting. Mol. Biotechnol. 201250237–249. [Google Scholar] [CrossRef]
- SharmaG.; ShenoyB.D. Multigene sequence-based identification of Colletotrichum cymbidiicolaC. karstii and C. phyllanthi from India. Czech Mycol. 20136579–88. [Google Scholar] [CrossRef]
- VieiraW.A.S.; NascimentoR.J.; MichereffS.J.; HydeK.D.; CâmaraM.P.S. First report of papaya fruit anthracnose caused by Colletotrichum brevisporum in Brazil. Plant Dis. 2013971659. [Google Scholar] [CrossRef] [PubMed]
- SainiT.J.; GuptaS.G.; AnandalakshmiR. First report of papaya anthracnose caused by Colletotrichum fructicola in India. New Dis. Rep. 2016347. [Google Scholar] [CrossRef]
- Tapia-TussellR.; Cortes-VelazquezA.; Valencia-YahT.; NavarroC.; EspinozaE.; MorenoB.; Perez-BritoD. First report of Colletotrichum magnum causing anthracnose in papaya in Mexico. Plant Dis. 20161002323. [Google Scholar] [CrossRef]
- SainiT.J.; GuptaS.G.; AnandalakshmiR. First report of papaya anthracnose caused by Colletotrichum salsolae in India. New Dis. Rep. 20173527. [Google Scholar] [CrossRef]
- Molina-ChavesA.; Gómez-AlpízarL.; Umaña-RojasG. Identificación de especies del género Colletotrichum asociadas a la antracnosis en papaya (Carica papaya L.) EnCosta Rica. Agron. Costarric. 20174169–80. [Google Scholar]
- DuanC.H.; PanH.R.; WangC.C. First report of Colletotrichum brevisporum causing anthracnose on papaya in Taiwan. Plant Dis. 20181022375–2376. [Google Scholar] [CrossRef]
- AktaruzzamanM.; AfrozT.; LeeY.G.; KimB.S. Post-harvest anthracnose of papaya caused by Colletotrichum truncatum in Korea. Eur. J. Plant Pathol. 2018150259–265. [Google Scholar] [CrossRef]
- SunY.C.; DammU.; HuangC.J. Colletotrichum plurivorumthe causal agent of anthracnose fruit rot of papaya in Taiwan. Plant Dis. 20191031040. [Google Scholar] [CrossRef]
- LiuX.B.; YanliF.; XiaolanZ.; HuangG.X. First report of papaya anthracnose caused by Colletotrichum brevisporum in China. Plant Dis. 20191032473. [Google Scholar] [CrossRef]
- DiasL.R.C.; BritoR.A.S.; MeloT.A.; SerraI.M.R.S. First report of papaya fruit anthracnose caused by Colletotrichum okinawense in Brazil. Plant Dis. 20204573. [Google Scholar] [CrossRef]
- SunY.C.; HuangC.J. Colletotrichum okinawensethe causal agent of postharvest anthracnose fruit rot of papaya in Taiwan. J. Plant Pathol. 2020102581–582. [Google Scholar] [CrossRef]
- ZhangY.; SunW.; NingP.; GuoT.; HuangS.P.; TangL.; QiliL.; MoJ. First report of anthracnose of papaya (Carica papaya L.) caused by Colletotrichum siamense in China. Plant Dis. 2021. [Google Scholar] [CrossRef]
- TabaS.; MikamiD.; TakaesuK.; OoshiroA.; MoromizatoZ.; NakasoneS.; KawanoS. Anthracnose of pitaya (Hylocereus undatus) by Colletotrichum gloeosporioides. Jpn. J. Phytopathol. 20067225–27. [Google Scholar] [CrossRef]
- PalmateerA.J.; PloetzR.C.; van SantenE.; CorrellJ.C. First occurrence of anthracnose caused by Colletotrichum gloeosporioides on Pitahaya. Plant Dis. 200791631. [Google Scholar] [CrossRef] [PubMed]
- TakahashiL.M.; RosaD.D.; BassetoM.A.; de SouzaH.G.; FurtadoE.L. First report of Colletotrichum gloeosporioides on Hylocereus megalanthus in Brazil. Australas. Plant Dis. 2008396–97. [Google Scholar] [CrossRef]
- MasyahitM.; KamaruzamanS.; YahyaA.; Mohd GhazaliM.S. The first report of the occurrence of anthracnose disease caused by (Penz.) Penz. & Sacc. on dragon fruit (Hylocereus spp.) in Peninsular Malaysia. Am. J. Appl. Sci. 20096902–912. [Google Scholar]
- MaW.J.; YangX.; WangX.R.; ZengY.S.; LiaoM.D.; ChenC.J.; SunS.; JiaD.M. First report of anthracnose disease on young stems of bawanghua (Hylocereus undatus) caused by Colletotrichum gloeosporioides in China. Plant Dis. 201498991. [Google Scholar] [CrossRef] [PubMed]
- GuoL.W.; WuY.X.; HoH.H.; SuY.Y.; MaoZ.C.; HeP.F.; HeY.Q. First report of dragon fruit (Hylocereus undatus) anthracnose caused by Colletotrichum truncatum in China. J. Phytopathol. 2014162272–275. [Google Scholar] [CrossRef]
- MeetumP.; LeksomboonC.; KanjanamaneesathianM. First report of Colletotrichum aenigma and C. siamensethe causal agents of anthracnose disease of dragon fruit in Thailand. J. Plant Pathol. 201597391–403. [Google Scholar]
- SuziantiI.V.; Intan SakinahM.A.; LatiffahZ. Characterization and Pathogenicity of Colletotrichum truncatum causing stem anthracnose of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. J. Phytopathol. 201516367–71. [Google Scholar]
- LinC.P.; AnnP.J.; HuangH.C.; ChangT.T.; TsaiJ.N. Anthracnose of pitaya (Hylocereus spp.) caused by Colletotrichum spp.a new postharvest disease in Taiwan. J. Taiwan Agric. Res. 201766171–183. [Google Scholar]
- ZhaoH.J.; ChenS.C.; ChenY.F.; ZouC.C.; WangX.L.; WangZ.H.; LiuA.R.; AhammedG.J. First report of red dragon fruit (Hylocereus polyrhizus) anthracnose caused by Colletotrichum siamense in China. Plant Dis. 20186862. [Google Scholar] [CrossRef]
- AbiramiK.; SakthivelK.; SheoranN.; BaskaranV.; GautamR.K.; JerardB.A.; KumarA. Occurrence of anthracnose disease caused by Colletotrichum siamense on dragon fruit (Hylocereus undatus) in Andaman IslandsIndia. Plant Dis. 2019103768. [Google Scholar] [CrossRef]
- NascimentoM.B.; BelléC.; AzambujaR.M.; MaichS.L.P.; NevesC.G.; Souza-JuniorI.T.; JacobsenC.R.F.; BarrosD.R. First Report of Colletotrichum karstii causing anthracnose spot on pitaya (Hylocereus undatus) in Brazil. Plant Dis. 20191032137. [Google Scholar] [CrossRef]
- OmarA.A.W. Occurrence of Colletotrichum anthracnose disease of guava fruit in Egypt. Int. J. Pest. Manag. 200147147–152. [Google Scholar]
- AmusaN.A.; AshayeO.A.; AmadiJ.; DapoO.O. Guava fruit anthracnose and the effects on its nutritional and market values in Ibadan. Niger. J. Appl. Sci. 20056539–542. [Google Scholar] [CrossRef]
- KeithL.M.; ZeeF.T. Guava disease in Hawaii and the characterization of Pestalotiopsis spp. affecting guava. Acta Hortic. 2010849269–276. [Google Scholar] [CrossRef]
- MeridaM.; PalmateerA.J. Florida Plant Disease Management Guide: Guava (Psidium guajava); PP-232. Plant Pathology DepartmentUF/IFAS Extension; Department of AgricultureCooperative Extension Service; Universidad de Florida: GainesvilleFLUSA2013. [Google Scholar]
- Intan SakinahM.A.; SuziantiI.V.; LatiffahZ. Molecular characterization and pathogenicity of Colletotrichum sp. from guava. Arch. Phytopathol. Plant Protect. 2014471549–1556. [Google Scholar]
- CruzA.F.; MedeirosN.L.; BenedetG.L.; AraújoM.B.; UesugiC.H.; da FerreiraM.A.S.V.; PeixotoJ.R.; BlumL.E.B. Control of post-harvest anthracnose infection in guava (Psidium guajava) fruits with phosphitescalcium chlorideacetyl salicylic acidhot waterand 1-MCP. Hort. Environ. Biotechnol. 201556330–340. [Google Scholar] [CrossRef]
- CrousP.W.; WingfieldM.J.; GuarroJ.; Hernández-Restrepo SuttonD.A.; AcharyaK.; BarberP.A.; BoekhoutT.; DimitrovR.A.; Dueñas DuttaA.K.; GenéJ.; et al. Fungal planet description sheets: 320–370. Persoonia 201534167–266. [Google Scholar] [CrossRef]
- SharmaG.; Kumar-PinnakaA.; ShenoyB.D. Resolving the Colletotrichum Siamese species complex using ApMat marker. Fungal Divers. 201571247–264. [Google Scholar] [CrossRef]
- BragançaC.A.D.; DammU.; BaroncelliR.; Massola JúniorN.S.; CrousP.W. Species of the Colletotrichum acutatum complex associated with anthracnose diseases of fruit in Brazil. Fungal Biol. 2016120547–561. [Google Scholar] [CrossRef] [PubMed]
- YaoJ.; LanC.; HuangP.; YuD. PCR detection of Colletotrichum gloeosporioides in Psidium guajava. Australas. Plant Pathol. 20184795–100. [Google Scholar] [CrossRef]
- NelsonS. Anthracnose of avocado. Plant Disease. 2008. PD-58. Available online: https://www.ctahr.hawaii.edu/oc/freepubs/pdf/pd-58.pdf (accessed on 15 April 2020).
- Silva-RojasH.V.; Ávila-QuezadaG.D. Phylogenetic and morphological identification of Colletotrichum boninense: A novel causal agent of anthracnose in avocado. Plant Pathol. 201160899–908. [Google Scholar] [CrossRef]
- HunupolagamaD.M.; WijesunderaR.L.C.; ChandrasekharanN.V.; WijesunderaW.S.S.; KathriarachchiH.S.; FernandoT.H.P.S. Characterization of Colletotrichum isolates causing avocado anthracnose and first report of C. gigasporum infecting avocado in Sri Lanka. Plant Pathol. Quar. 20155132–143. [Google Scholar] [CrossRef]
- Hernández-LauzardoA.N.; Campos-MartínezA.; Velázquez-del ValleM.G.; Flores-MoctezumaH.E.; Suárez-RodríguezR.; Ramírez-TrujilloJ.A. First report of Colletotrichum godetiae causing anthracnose on avocado in Mexico. Plant Dis. 201599555. [Google Scholar] [CrossRef]
- AkgulD.S.; AwanQ.N.; GulerP.G.; ÖnelgeN. First report of anthracnose and stem end rot diseases caused by Colletotrichum gloeosporioides and Neofusicoccum australe on avocado fruits in Turkey. Plant Dis. 20161001792. [Google Scholar] [CrossRef]
- ShivasR.G.; TanY.P.; EdwardsJ.; DinhQ.; MaxwellA.; AndjicV.; LiberatoJ.R.; AndersonC.; BeasleyD.R.; BransgroveK.; et al. Colletotrichum species in Australia. Australas. Plant Pathol. 201645447–464. [Google Scholar] [CrossRef]
- HongerJ.O.; OffeiS.K.; OduroK.A.; OdamttenG.T.; NyakuS.T. Identification and molecular characterisation of Colletotrichum species from avocadocitrus and pawpaw in Ghana. S. Afr. J. Plant Soil 2016331–9. [Google Scholar] [CrossRef]
- SharmaG.; MaymonM.; FreemanS. Epidemiologypathology and identification of Colletotrichum including a novel species associated with avocado (Persea americana) anthracnose in Israel. Sci. Rep. 2017715839. [Google Scholar] [CrossRef]
- Fuentes-AragónD.; Juárez-VázquezS.B.; Vargas-HernándezM.; Silva-RojasH.V. Colletotrichum fructicolaa member of Colletotrichum gloeosporioides sensu latois the causal agent of anthracnose and soft rot in avocado ruits cv. “Hass”. Mycobiology 20184692–100. [Google Scholar] [CrossRef]
- GiblinF.R.; TanY.P.; MitchellR.; CoatesL.M.; IrwinJ.A.G.; ShivasR.G. Colletotrichum species associated with pre-and post-harvest diseases of avocado and mango in eastern Australia. Australas. Plant Pathol. 201847269–276. [Google Scholar] [CrossRef]
- KwonJ.-H.; ChoiO.; LeeY.; KimS.; KangB.; KimJ. Anthracnose on postharvest avocado caused by Colletotrichum kahawae subsp. ciggaro in South Korea. Can. J. Plant Pathol. 202042508–513. [Google Scholar] [CrossRef]
- Ayvar-SernaS.; Díaz-NájeraJ.F.; MateoV.H.; Camacho-TapiaM.; Valencia-RojasG.A.; LimaN.B.; Tovar-PedrazaJ.M. First report of Colletotrichum jiangxiense causing avocado anthracnose in Mexico. Plant Dis. 2021105502. [Google Scholar] [CrossRef]
- Fuentes-AragónD.; Silva-RojasH.V.; GuarnacciaV.; Mora-AguileraJ.A.; Aranda-OcampoS.; Bautista-MartínezN.; Téliz-OrtízD. Colletotrichum species causing anthracnose on avocado fruit in Mexico: Current status. Plant Pathol. 2020691513–1528. [Google Scholar] [CrossRef]
- SoaresM.G.O.; AlvesE.; SilveiraA.L.; PereiraF.D.; GuimarãesS.S.C. Colletotrichum siamense is the main aetiological agent of anthracnose of avocado in south-eastern Brazil. Plant Pathol. 202170154–166. [Google Scholar] [CrossRef]
- Bananalink. All about Banana. 2020. Available online: http://www.bananalink.org.uk/all-about-bananas (accessed on 21 May 2020).
- Agriexchange. Banana Report. 2020. Available online: https://agriexchange.apeda.gov.in/Weekly_eReport/Banana_Report.pdf (accessed on 21 May 2020).
- Food and Agriculture Organization of the United Nations (FAO). Banana Market Review February 2020 Snapshot; FAO: RomeItaly2020. [Google Scholar]
- JegerM.J.; Eden-GreenS.; ThreshJ.M.; JohannsonA.; WallerJ.M.; BrownA.E. Banana diseases. In Banana and Plantains; GowenS.R.Ed.; Chapman and Hall: LondonUK1995; pp. 3117–3381. [Google Scholar]
- MeredithD.S. Studies on Gloeosporium musarum (Gke and massee) causing storage rots of Jamaican bananas. Anthracnose and its chemical control. Ann. Appl. Biol. 196048279–290. [Google Scholar] [CrossRef]
- ChilletM.O.; HubertL.; de BellaireL.D.L. Relationship between physiological ageripening and susceptibility of banana to wound anthracnose. Crop. Prot. 2007261078–1082. [Google Scholar] [CrossRef]
- Abd-ElsalamK.A.; RoshdyS.; AminO.E.; RabaniM. First morphogenetic identication of the fungal pathogen Colletotrichum musae (Phyllachoraceae) from imported bananas in Saudi Arabia. Genet. Mol. Res. 201092335–2342. [Google Scholar] [CrossRef] [PubMed]
- SuY.Y.; NoireungP.; LiuF.; HydeK.D.; MoslenM.A.; BahkaliA.H.; Abd-ElsalamK.A.; CaiL. Epitypification of Colletotrichum musaethe causative agent of banana anthracnose. Mycoscience 201152376–382. [Google Scholar] [CrossRef]
- UdayangaD.; ManamgodaD.S.; LiuX.Z.; ChukeatiroteE.; HydeK.D. What are the common anthracnose pathogens of tropical fruits? Fungal Divers. 201361165–179. [Google Scholar] [CrossRef]
- BalendresM.A.; MendozaJ.; Dela CuevaF. Characteristics of Colletotrichum musae PHBN0002 and the susceptibility of popular banana cultivars to postharvest anthracnose. Indian Phytopathol. 20207357–64. [Google Scholar] [CrossRef]
- SangeethaG.; UsharaniS.; MuthukumarA. Significance of Lasiodiplodia theobromae and Colletotrichum musae in causing crown rot in banana and their reaction on some commercial banana cultivars. Indian J. Hort. 20106721–25. [Google Scholar]
- PatilR.S.; DeshmukhR.G.; BhaskarK.R.; JahagirdarS.W. Growth and export performance of mango in India. Int. J. Curr. Microbiol. Appl. Sci. 201862667–2673. [Google Scholar]
- ArauzL.F. Mango anthracnose: Economic impact and current options for integrated management. Plant Dis. 200084600–611. [Google Scholar] [CrossRef] [PubMed]
- ChowdappaP.; Mohan KumarS.P. Existence of two genetically distinct populations of Colletotrichum gloeosporioides Penz in mango from India. Pest. Manag. Hort. Ecosyst. 201218161–170. [Google Scholar]
- JayasingheC.K.; FernandoT.H.P.S. First report of Colletotrichum acutatum on Mangifera indica in Sri Lanka. J. Sci. Bio. Sci. 20093831–34. [Google Scholar] [CrossRef]
- KamleM.; KumarP.; GuptaV.K.; TiwariA.K.; MisraA.K.; PandeyB.K. Identification and phylogenetic correlation among Colletotrichum gloeosporioides pathogen of anthracnose for mango. Biocatal. Agric. Biotechnol. 20132285–287. [Google Scholar] [CrossRef]
- AlahakoonP.W.; BrownA.E.; SreenivasaprasadS. Cross-infection potential of genetic groups of Colletotrichum gloeospoirides on tropical fruits. Physiol. Mol. Plant. Pathol. 19944493–103. [Google Scholar] [CrossRef]
- HaydenH.L.; PeggK.G.; AitkenE.A.B.; IrwinJ.A.G. Genetic relationships as assessed bimolecular markers and cross- infection among strains of Colletotrichum gloeosporioides. Aust. J. Bot. 1994429–18. [Google Scholar] [CrossRef]
- WallerJ.M.; BridgeP.D. Recent advances in understanding Colletotrichum diseases of some tropical perennial crops. In Colletotrichum: Host SpecificityPathologyand Host-Pathogen Interaction; PruskyD.FreemanS.DickmanM.B.Eds.; APS Press: St PaulMNUSA2000; pp. 337–345. [Google Scholar]
- PloetzR. Anthracnose: The most important disease in much of the mango-producing world. In PLP NewsThe Newsletter of the Plant Pathology Department; The University of Florida: GainsevilleFLUSA1999; Volume 3pp. 1–2. [Google Scholar]
- LimaW.G.; SpositoM.B.; AmorimL.; GolcalvesF.P.; de FilhoP.A.M. Comparative epidemiology of Colletotrichum species from mango in northeastern Brazil. Eur. J. Plant Pathol. 2015141679–688. [Google Scholar] [CrossRef]
- SharmaG.; GryzenhoutM.; HydeK.D.; PinnakaA.K.; ShenoyB.D. First report of Colletotrichum asianum causing mango anthracnose in South Africa. Plant Dis. 201599725. [Google Scholar] [CrossRef]
- BenatarG.V.; WibowoA.; Suryant. First report of Colletotrichum asianum associated with mango fruit anthracnose in Indonesia. Crop. Prot. 2021141105432. [Google Scholar] [CrossRef]
- FAOSTAT. 2017. Available online: http://www.fao.org/fileadmin/templates/est/COMM_MARKETS_MONITORING/Tropical_Fruits/Documents/CA2895EN (accessed on 15 April 2020).
- DickmanK.B.; PartV. Papaya: Anthracnose. In Compendium of Tropical Fruit Diseases; PloetzR.C.ZentmeyerG.A.NishijimaW.T.RohrbachK.G.OhrH.D.Eds.; APS Press: St PaulMNUSA1994; pp. 58–59. [Google Scholar]
- Vieria dos SantosW.A.; dos Santos NunesA.; VelosoJ.S.; MachadoA.R.; Queiroz BalbinoV.; da SilvaA.C.; Medeiros GomesA.Â.; DoyleV.P.; CâmaraM.P.S. Colletotrichum truncatum causing anthracnose on papaya fruit (Carica papaya) in Brazil. Australas. Plant Dis. Notes 202015. [Google Scholar] [CrossRef]
- Pérez-BritoD.; Cortes-VelázquezA.; Valencia-YahT.; Magaña-ÁlvarezA.; NavarroC. Genetic variation of Colletotrichum magnum isolated from Carica papaya as revealed by DNA fingerprinting. J. Microbiol. 20186813–821. [Google Scholar] [CrossRef]
- SekeliR.; HamidM.H.; RazakR.A.; WeeC.Y.; Ong-AbdullahJ. Malaysian Carica papaya L. var. Eksotika: Current research strategies fronting challenges. Front. Plant Sci. 201891380. [Google Scholar] [CrossRef]
- ChenN.J.; PaullR.E. Overall Dragon Fruit Production and Global Marketing. FFTC Agricultural Policy Platform (FFTC-AP). 2018. Available online: https://ap.fftc.org.tw/article/1596 (accessed on 15 April 2020).
- YacobA. Dragonfruit: The Next Potential Major Tropical Fruit? 2019. Available online: https://www.itfnet.org/v1/2019/10/dragonfruit-the-next-potential-major-tropical-fruit/ (accessed on 15 April 2020).
- HoaT.T.; ClarkC.J.; WaddellB.C.; WoolfA.B. Postharvest quality of dragon fruit (Hylocereus undatus) following disinfesting hot air treatments. Postharvest Biol. Technol. 20064162–69. [Google Scholar] [CrossRef]
- GunasenaH.P.M.; PushpakumaraD.K.N.G.; KariyawasamM. Dragon fruit (Hylocerus undatus [Haw.] Britton and Rose). In Underutilized Fruit Trees in Sri Lanka; PushpakumaraD.K.N.G.GunasenaH.P.M.SinghV.P.Eds.; South Asia Regional Office; World Agroforestry Centre (International Council for Research in Agroforestry–ICRAF): New DelhiIndia2007; Volume 1pp. 110–141. [Google Scholar]
- LeiteK.M.S.C.; TadiottiA.C.; BaldochiD.; OliveiraO.M.M.F. Partial purificationheat stability and kinetic characterization of the pectinmethylesterase from Brazilian guavaPaluma cultivars. Food Chem. 200694565–572. [Google Scholar] [CrossRef]
- NaseerS.; NaureenN.; HussainS.; PervaizM.; RahmanM. The phytochemistry and medicinal value of Psidium guajava (guava). Clin. Phytosci. 2018432. [Google Scholar] [CrossRef]
- MortonJ. Guava (Psidium guajava L.). In Fruits of Warm Climates; Morton: MiamiFLUSA1987; pp. 356–363. Available online: http://www.hort.purdue.edu/newcrop/morton/guava.html# (accessed on 20 April 2020).
- ParionaA. Top Guava Producing Countries in the World. 2017. Available online: https://www.worldatlas.com/articles/top-guava-producing-countries-in-the-world.html (accessed on 20 May 2020).
- DasM.; BoraK.N. Ultrastructural studies on infection processes by Colletotrichum acutatum on guava fruit. Indian Phytopathol. 199851353–356. [Google Scholar]
- PeresN.A.; TimmerL.W.; AdaskavegJ.; CorrellJ.C. Life cycles of Colletotrichum acutatum. Plant Dis. 200589784–796. [Google Scholar] [CrossRef] [PubMed]
- SoaresA.R.; LourencoS.; AmorimL. Infection of guava by C. gleosporioides and C. acutatum under different temperature and wetting periods. Trop. Plant Pathol. 200833265–272. [Google Scholar]
- WorkmanD. Avocados Export by Country. 2020. Available online: http://www.worldstopexports.com/avocados-exports-by-country/ (accessed on 1 July 2020).
- NgboluaK.; NgialaG.B.; LiyongoC.I.; AshandeC.M.; LufuluaboG.L.; MukizaJ.; MpianaP.T. A mini review on the phytochemistry and pharmacology of the medicinal plant species Persea americana Mill. (Lauraceae). Discov. Phytomed. 20196102–111. [Google Scholar] [CrossRef]
- Velázquez-del ValleM.G.; Campos-MartínezA.; Flores-MoctezumaH.E.; Suárez-RodríguezR.; Ramírez-TrujilloJ.A.; Hernández-LauzardoA.N. First report of avocado anthracnose caused by Colletotrichum karstii in Mexico. Plant Dis. 2016100534. [Google Scholar] [CrossRef]
- EckertJ.W. Recent developments in the chemical control of postharvest diseases. Acta Hortic. 1990269477–494. [Google Scholar] [CrossRef]
- CoatesL.M.; JohnsonG.I.; DaleM. Postharvest pathology of fruit and vegetables. In Plant Pathogens and Plant Diseases; BrownJ.OgleH.Eds.; Rockvale Publications: BurlingtonMAUSA1997; pp. 533–547. [Google Scholar]
- NorthoverJ.; ZhouT. Control of rhizopus rot of peaches with postharvest treatments of tebuconazolefludioxoniland Pseudomonas syringae. Can. J. Plant Pathol. 200224144–153. [Google Scholar] [CrossRef]
- SharmaR.R.; SinghD.; SinghR. Biological control of postharvest diseases of fruits and vegetables by microbial antagonists: A review. Biol. Control. 200950205–221. [Google Scholar] [CrossRef]
- WisniewskiM.; WilsonC.; DrobyS.; ChalutzE.; El GhaouthA.; StevensC. Postharvest biocontrol: New concepts and applications. In Biological Control: A Global Perspective; Vincent.C.GoettalM.S.LazarovitsG.Eds.; CABI: CambridgeMAUSA2007; pp. 262–273. [Google Scholar]
- SpadaroD.; DrobyS. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 20164739–49. [Google Scholar] [CrossRef]
- FarboM.G.; UrgegheP.P.; FioriS.; MarcelloA.; OggianoS.; BalmasV.; HassanZ.U.; JaouaS.; MigheliQ. Effect of yeast volatile organic compounds on ochratoxin A-producing Aspergillus carbonarius and A. ochraceus. Int. J. Food Microbiol. 20182841–10. [Google Scholar] [CrossRef]
- AsgarA.; NoosheenZ.; SivakumarM.; YasmeenS.; AldersonP.G.; MehdiM. Effectiveness of submicron chitosan dispersions in controlling anthracnose and maintaining quality of dragon fruit. Postharvest Biol. Technol. 201386147–153. [Google Scholar]
- De OliveiraK.A.R.; BergerL.R.R.; de AraújoS.A.; CâmaraM.P.S.; de SouzaE.L. Synergistic mixtures of chitosan and Mentha piperita L. essential oil to inhibit Colletotrichum species and anthracnose development in mango cultivar Tommy Atkins. Food Microbiol. 20176696–103. [Google Scholar] [CrossRef] [PubMed]
- BragaS.; LundgrenG.; SamaraM.; Tavares JoseanT.; Willie AndersonV.; CâmaraM.; EvandroS. Application of coatings formed by chitosan and Mentha essential oils to control anthracnose caused by Colletotrichum gloesporioides and C. brevisporum in papaya (Carica papaya L.) fruit. Int. J. Biol. Macromol. 2019139631–639. [Google Scholar] [CrossRef]
- PalouL. Postharvest treatments with GRAS salts to control fresh fruit decay. Horticulturae 2018446. [Google Scholar] [CrossRef]
- De CostaD.M.; GunawardhanaH.M.D.M. Effects of sodium bicarbonate on pathogenicity of Colletotrichum musae and potential for controlling postharvest diseases of banana. Postharvest Biol. Technol. 20126854–63. [Google Scholar] [CrossRef]
- JitareeratP.; SripongK.; MasayaK.; AiamlaorS.; UthairatanakijA. Combined effects of food additives and heat treatment on fruit rot disease and quality of harvested dragon fruit. Agric. Nat. Resour. 20185543–549. [Google Scholar] [CrossRef]
- KalupahanaK.I.M.; KuruppuM.; DissanayakeP.K. Effect of essential oils and GRAS compounds on postharvest disease control in mango (Mangifera indica L.cv Tom EJC). J. Agric. Sci. Sri Lanka 202015207–221. [Google Scholar] [CrossRef]
- De la Rosa-GarcíaS.; Martínez-TorresP.; Gomez-CornelioS.; Corral-AguadoM.; QuintanaP.; Gómez-OrtízN. Antifungal activity of ZnO and MgO nanomaterials and their mixtures against Colletotrichum gloeosporioides strains from tropical fruit. J. Nanomater. 20183498527. [Google Scholar] [CrossRef]
- JaganaD.; HegdeY.R.; LellaR. Green nanoparticles: A novel approach for the management of banana anthracnose caused by Colletotrichum musae. Int. J. Curr. Microbiol. Appl. Sci. 201761749–1756. [Google Scholar] [CrossRef]
- ChowdappaP.; GowdaS.; ChethanaC.S.; MadhuraS. Antifungal activity of chitosan-silver nanoparticle composite against Colletotrichum gloeosporoides associated with mango anthracnose. Afr. J. Microbiol. Res. 201481803–1812. [Google Scholar]
- RaghavendraS.N.; RaghuH.S.; DivyashreeK.; RajeshwaraA.N. Antifungal efficiency of copper oxychloride-conjugated silver nanoparticles against Colletotrichum gloeosporioides which causes anthracnose disease. Asian J. Pharm. Clin. Res. 201912230–233. [Google Scholar] [CrossRef]
- TurnerM. Physics of cold plasma. In Cold Plasma in Food and Agriculture; MisraN.N.SchluterO.CullenP.J.Eds.; Academic Press: San DiegoCAUSA2016; pp. 17–51. [Google Scholar]
- MisraN.N.; BarunY.; RoopeshM.S.; CheorunJ. Cold plasma for effective fungal and mycotoxin control in Foods: Mechanismsinactivation effects and applications. Compr. Rev. Food Sci. Saf. 201918106–120. [Google Scholar] [CrossRef]
- SiddiqueS.S.; HardyG.J.; BaylissK.L. Advanced technologies for controlling postharvest diseases of fruit. Acta Hortic. 20181194193–200. [Google Scholar] [CrossRef]
- MisraN.N.; MoiseevT.; PatilS.; PankajS.K.; BourkeP.; MosnierJ.P.; KeenerK.M.; CullenP.J. Cold plasma in modified atmospheres for post-harvest treatment of strawberries. Food Bioprocess Tech. 201473045–3054. [Google Scholar] [CrossRef]
- TalhinhasP.; GonçalvesE.; SreenivasaprasadS.; OliveiraH. Virulence diversity of anthracnose pathogens (Colletotrichum acutatum and C. gloeosporioides Species Complexes) on eight olive cultivars commonly grown in Portugal. Eur. J. Plant Pathol. 201414273–83. [Google Scholar] [CrossRef]
- BaroncelliR.; ZapparataA.; SarroccoS.; SuknoS.A.; LaneC.R.; ThonM.R.; VannacciG.; HolubE.; SreenivasaprasadS. Molecular diversity of anthracnose pathogen populations associated with UK strawberry production suggests multiple introductions of three different Colletotrichum species. PLoS ONE 201510e0129140. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fruit Crop | Reported Colletotrichum spp. | Country | References |
|---|---|---|---|
| Banana (Musa spp.) | C. paxtonii | St Lucia | Damm et al. (2012a) [12] |
| C. gloeosporioides sensu lato | Malaysia | Intan Sakinah et al. (2013) [20] | |
| C. scovillei | Hainan ProvinceChina | Zhou et al. (2016) [21] | |
| C. siamense | India | Kumar et al. (2017) [22] | |
| C. siamenseC. tropicaleC. chrysophilumC. theobromicola | Brazil | Vieira et al. (2017) [23] | |
| C. gloeosporioides | Ecuador | Riera et al. (2019) [24] | |
| C. siamense | Turkey | Uysal and Kurt (2020) [25] | |
| C. chrysophilum | Mexico | Fuentes-Aragón et al. (2020) [26] | |
| Mango (Mangifera indica L.) | C. asianum and C. gloeosporioides | Panama | Rojas et al. (2010) [27] |
| C. asianumC. fructicolaC. tropicaleC. karstii, C. dianesei | northeastern Brazil | Lima et al. (2013) [28] | |
| C. fragariae sensu strictoC. fructicolaC. jasmine-sambac, C. melanocaulon | India | Sharma et al. (2013) [29] | |
| C. asianumC. gloeosporioides | Sri Lanka. | Krishnapillai and Wijeratnam (2014) [30] | |
| C. asianum | Malaysia | Latiffah et al. (2015) [31] | |
| C. asianumC. gloeosporioides | Colombia | Pardo-De la Hoz et al. (2016) [32] | |
| C. asianum | Sanya CityChina | Qin et al. (2017) [33] | |
| C. asianumC. fructicolaC. siamense | GuangxiSouth China | Mo et al. (2018) [34] | |
| C. scovillei (mango leaves) | GuangxiChina | Qin et al. (2017) [33] | |
| C. asianumC. cliviicolaC. cordylinicolaC. endophytica, C. fructicolaC. gigasporumC. gloeosporioidesC. karstii, C. liaoningenseC. musaeC. scovilleiC. siamense, C. tropicale | southern China | Li et al. (2019) [35] | |
| C. asianumC. fructicolaC. siamenseC. tropicaleC. scovillei | Taiwan | Wu et al. (2020) [36] | |
| C. alienumC. asianumC. fructicolaC. siamense, C. tropicale | Mexico | Tovar-Pedraza et al. (2020) [37] | |
| C. asianum | Philippine | Alvarez et al. (2020) [38] | |
| C. alienum | BeijingChina | Ahmad et al. (2021) [39] | |
| Papaya (Carica papaya L.) | C. gloeosporioidesC. capsici | Yucatan peninsulaMexico | Tapia-Tussell et al. (2008) [40] |
| C. gloeosporioidesC. capsici | Malaysia | Rahman et al. (2008) [41] | |
| C. gloeosporioidesC. capsici | Miyako IslandsOkinawaJapan | Yaguchi et al. (2008) [42] | |
| C. gloeosporioidesC. capsici | South Florida | Tarnowski and Ploetz (2010) [43] | |
| C. magnaC. gloeosporioides | Brazil | Nascimento et al. (2010) [44] | |
| C. gloeosporioidesC. capsiciC. dematium | YucatanMexico | Santamaría Basulto et al. (2017) [45] | |
| C. acutatumC. simmondsii | Australia | Damm et al. (2012a) [12] | |
| C. karstii | Brazil | Damm et al. (2012b) [46] | |
| C. queenslandicum | Australia | Weir et al. (2012) [13] | |
| C. siamense | South Africa | Weir et al. (2012) [13] | |
| C. gloeosporioidesC. capsici | Trinidad Island | Rampersad (2011) [47] Maharaj and Rampersad (2013) [48] | |
| C. karstii | India | Sharma and Shenoy (2013) [49] | |
| C. brevisporum | Brazil | Vieira et al. (2013) [50] | |
| C. fructicolaC. gloeosporioides | India | Saini et al. (2016) [51] | |
| C. magnum | Mexico | Tapia-Tussell et al. (2016) [52] | |
| C. salsolae | India | Saini et al. (2017) [53] | |
| C. truncatumC. gloeosporioides sensu latoC. magnum | Costa Rica | Molina-Chaves et al. (2017) [54] | |
| C. brevisporum | Taiwan | Duan et al. (2018) [55] | |
| C. truncatum | Korea | Aktaruzzaman et al. (2018) [56] | |
| C. plurivorum | Taiwan | Sun et al. (2019) [57] | |
| C. brevisporum | China | Liu et al. (2019) [58] | |
| C. truncatum | Brazil | Vieira et al. (2017) [23] | |
| C. okinawense | Brazil | Dias et al. (2020) [59] | |
| C. okinawense | Taiwan | Sun and Huang (2020) [60] | |
| C. siamense | China | Zhang et al. (2021) [61] | |
| Dragon fruits (Hylocereus spp.) | C. gloeosporioides sensu lato (H. undatus) | Okinawa PrefectureJapan | Taba et al. (2006) [62] |
| C. gloeosporioides sensu lato (H. undatus) | Miami-Dade CountyFloridaUSA | Palmateer et al. (2007) [63] | |
| C. gloeosporioides sensu lato (H. megalanthus) | Brazil | Takahashi et al. (2008) [64] | |
| C. gloeosporioides sensu lato (H. polyrhizusH. undatusSelenicereus megalanthus) | Malaysia | Masyahit et al. (2009) [65] | |
| C. gloeosporioides (H. undatus young stems) | China | Ma et al. (2014) [66] | |
| C. truncatum (H. undatus fruits) | Yuanjiang CountyYunnan ProvinceChina | Guo et al. (2014) [67] | |
| C. aenigma and C. siamense (H. undatus stem and fruit) | Thailand | Meetum et al. (2015) [68] | |
| C. truncatum (H. polyrhizus stem) | Malaysia | Suzianti et al. (2015) [69] | |
| C. gloeosporioidesC. truncatumC. boninense (H. polyrhizus, H. undatusH. costaricensis) | Taiwan | Lin et al. (2017) [70] | |
| C. siamense (H. polyrhizus stem) | China | Zhao et al. (2018) [71] | |
| C. siamense (H. undatus) | Andaman IslandsIndia | Abirami et al. (2019) [72] | |
| C. karstii (H. undatus stem) | Brazil | Nascimento et al. (2019) [73] | |
| Guava (Psidium guajava L.) | C. gloeosporioides sensu lato | Egypt | Omar (2001) [74] |
| C. gloeosporioides sensu lato | IbadanNigeria | Amusa et al. (2005) [75] | |
| C. gloeosporioides | Hawaii | Keith and Zee (2010) [76] | |
| C. psidii | Italy | Weir et al. (2012) [13] | |
| C. guajavae | India | Damm et al. (2012a) [12] | |
| C. gloeosporioides sensu lato | FloridaUSA | Merida and Palmateer (2013) [77] | |
| C. gloeosporioides sensu lato | Malaysia | Intan Sakinah et al. (2014) [78] | |
| C. simmondsii | Brazil | Cruz et al. (2015) [79] | |
| C. abscissum | USA | Crous et al. (2015) [80] | |
| C. siamense | India | Sharma et al. (2015b) [81] | |
| C. abscissum | Brazil | Bragança et al. (2016) [82] | |
| C. gloeosporioides sensu lato | China | Yao et al. (2018) [83] | |
| Avocado (Persea americana Mill.) | C. gloeosporioides sensu lato | USA | Nelson (2008) [84] |
| C. gloeosporioidesC. acutatumC. boninense | Mexico | Silva-Rojas and Avila-Quezada (2011) [85] | |
| C. gigasporum | Sri Lanka | Hunupolagama et al. (2015) [86] | |
| C. godetiae | Mexico | Hernandez-Lauzardo et al. (2015) [87] | |
| C. gloeosporioides | Mersin ProvinceTurkey | Akgul et al. (2016) [88] | |
| C. alienumC. fructicolaC. siamense | Australia | Shivas et al. (2016) [89] | |
| C. gloeosporioidesC. siamense | Ghana | Honger et al. (2016) [90] | |
| C. aenigmaC. alienumC. fructicolaC. gloeosporioides sensu strictoC. karstiiC. nupharicolaC. siamenseC. theobromicolaC. perseae | Israel | Sharma et al. (2017) [91] | |
| C. fructicola | HidalgoMexico | Fuentes-Aragón et al. (2018) [92] | |
| C. alienumC. asianumC. fructicolaC. karstiiC. siamense | eastern Australia | Giblin et al. (2018) [93] | |
| C. kahawae subsp. cigarro | JinjuSouth Korea | Kwon et al. (2020) [94] | |
| C. karstii | Turkey | Uysal andKurt (2020) [25] | |
| C. jiangxiense | Mexico | Ayvar-Serna et al. (2020) [95] | |
| C. karstiiC. godetiaeC. siamenseC. fioriniaeC. cigarroC. chrysophilumC. jiangxienseC. tropicaleC. nymphaeae,Colletotrichum sp. 1Colletotrichum sp. 2 | Mexico | Fuentes-Aragón et al. (2020) [96] | |
| C. siamenseC. kartsii | south eastern Brazil | Soares et al. (2021) [97] |
| Fruit Crop | Colletotrichum spp. | Infected Parts |
|---|---|---|
| Banana (Musa spp.) | C. scovillei,C. gloeosporioides,C. siamense, C. tropicaleC. chrysophilumC. paxtonii C. theobromicola | Fruit |
| C. musae | Ripe fruitbut has been reported from leaves and root of Musa spp. | |
| Mango (Mangifera indica L.) | C. fructicolaC. tropicaleC. fragariae sensu strictoC. jasmine-sambacC. melanocaulon, C. alienum | Fruit |
| C. asianumC. karstiiC. scovilleiC. scovillei, C. fructicolaC. siamenseC. cliviicola, C. musae C. cordylinicolaC. endophytica, C. gigasporumC. liaoningenseC. tropicale | Fruit and leaves | |
| C. gloeosporioides | Fruitleavesand inflorescence | |
| Papaya (Carica papaya L.) | C. magnaC. gloeosporioidesC. dematium,
C. acutatumC. simmondsiiC. karstii, C. queenslandicumC. siamenseC. salsolae, C. magnumC. brevisporumC. fructicola, C. plurivorumC. okinawenseC. siamense | Fruit |
| Dragon fruits (Hylocereus spp.) | C. gloeosporioides sensu latoC. truncatum, C. aenigmaC. siamense | Fruit and stem |
| C. gloeosporioides | Young stem | |
| C. boninense | Fruit | |
| C. karstii | Stem | |
| Guava (Psidium guajava L.) | C. gloeosporioides sensu latoC. psidii, C. guajavaeC. simmondsiiC. abscissum, C. siamense | Fruit |
| C. gloeosporioides | Fruit and leaves | |
| Avocado (Persea americana Mill.) | C. gloeosporioides sensu latoC. nymphaeae C. gloeosporioidesC. gigasporumC. karstii, C. godetiaeC. alienumC. fructicola, C. siamenseC. aenigmaC. alienum C. perseae C. nupharicolaC. theobromicolaC. tropicale. C. kahawae subsp. cigarroC. jiangxiense, C. cigarroC. chrysophilum | Fruit |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPIBaselSwitzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
ZakariaL. Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review. Agriculture 202111297. https://doi.org/10.3390/agriculture11040297
Zakaria L. Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review. Agriculture. 2021; 11(4):297. https://doi.org/10.3390/agriculture11040297
Chicago/Turabian StyleZakariaLatiffah. 2021. "Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review" Agriculture 11no. 4: 297. https://doi.org/10.3390/agriculture11040297
APA StyleZakariaL. (2021). Diversity of Colletotrichum Species Associated with Anthracnose Disease in Tropical Fruit Crops—A Review. Agriculture11(4)297. https://doi.org/10.3390/agriculture11040297